
2-3 유전
2-3-1 변이
(1) 변이의 종류
개체들 사이에 형질(形質)의 특성이 다른 것을 변이(變異; variation)라고 한다. 형질의 변이는 유전적 원인으로 나타나는 유전변이(遺傳變異; genetic variation)와 환경요인에 의한 환경변이(環境變異; environmental variation)가 있다. 유전변이는 다음 세대로 유전되지만 환경변이는 유전되지 않는다. 유전변이가 생기는 것은 감수분열 과정에서 일어나는 유전자재조합과 염색체와 유전자의 돌연변이가 주된 원인이다. 유전변이가 크다는 것은 유전자형이 다양하다는 것과 같은 의미이다.
유전변이는 형질의 특성에 따라 형태적 변이(키가 큰 것과 작은 것)와 생리적 변이(병해충에 강한 것과 약한 것)로 나눌 수 있다. 또한, 변이양상에 의하여 불연속변이(不連續變異 : 꽂 색깔이 붉은 것과 휜 것으로 뚜렷이 구별 되는 것)와 연속변이(連繽變異 : 키가 작은 것부터 큰 것에 이르기까지 여러 등급으로 나타나는 것)로 구분하며, 불연속변이를 하는 형질을 질적 형질 (質的形質; qualitative character), 연속변이를 하는 형질을 양적 형질(量的 形質; quantitative character) 이라고 한다.
질적 형질은 불연속변이를 하므로 표현형들의 구별이 쉽다. 따라서, 각 표현형에 속하는 개체수나 비율을 조사하여 유전분석을 하며, 원하는 유전자형을 쉽게 선발할 수 있다. 그러나 연속변이를 하는 양적 형질은 표현형의 구별이 어렵기 때문에 평균 • 분산 • 회귀 • 유전력 등 통계적 방법에 의하여 유전분석을 하고 그 결과를 선발에 이용한다.
(2) 변이의 작성
작물육종은 형질을 개량하기 위하여 자연변이를 이용하거나 인위적으로 변이를 작성하고 그 변이 중에서 원하는 유전자형의 개체를 선발하여 품종을 육성한다. 유전변이를 작성하는 방법에는 인공교배• 돌연변이유발• 염색체조작• 유전자전환등이 있다.
특성이 서로 다른 자방친과 화분친을 인공교배하면 양친의 대립유전자들이 새롭게 조합되므로 잡종 후대에 여러 종류의 유전자형이 분리하여 유전변이가 나타난다. 인공교배하는 양친의 유전적 차이가 클수록 잡종집단의 유전변이(유전자형의 다양성)가 커진다.
자연돌연변이의 발생빈도는 매우 낮다. 그래서 방사선이나 화학물질을 처리하여 인위적으로 돌연변이를 유발시킨다. 인위돌연변이는 인공교배처럼 여러 대립유전자들이 재조합되는 것이 아니어서 특정한 형질만 개량되는 특징이 있다.
염색체의 수와 구조가 변화하면 식물체는 형태적 및 생리적으로 다른 특 성을 나타낸다. 따라서 염색체를 인위적으로 조작하면 반수체 • 배수체 • 이수체 등 유전변이가 생기게 된다.
인공교배 • 인위돌연변이 및 염색체조작은 주로 같은 종 내에서 유전변이를 작성하는 방법이다. 그런데 세포융합(細胞融合; cell fusion)이나 유전자전환(遺傳子轉換; gene transformation) 기법을 이용하면 다른 종의 우량유전자를 도입한 유전변이를 만들 수 있다. 세포융합은 인공교배가 안되는 원연 종 • 속간에 유전자를 교환할 수 있는 방법이고 유전자전환은 생물종에 관계없이 원하는 유전자만을 도입할 수 있는 방법이다.
(3) 변이의 선발
작물육종에서는 우량한 변이를 선발하기 위해 형질의 특성검정(特性檢定) 을 한다. 식별이 간단하고 표현형으로 유전자형을 판정하기 쉬운 형질은 특별한 선발기술이 없어도 된다. 그러나 내병성이나 내냉성처럼 특정 환경에 서 발현하는 형질은 특성검정이 필요하다. 특성검정은 자연조건 • 검정포 • 실내 등을 이용한다. 형질의 특성을 검정하는 데는 인력 • 경비 • 시간 등이 많이 요구된다.
우량한 변이체를 고를 때 형질간의 상관관계(相關關係)를 이용하면 목표 형질을 선발하기 쉽다. 예컨대, 밀은 어린 식물의 잎색깔이 질은 것일수록 내한성이 강하고 콩의 단백질 함량은 비중과 높은 정상관이 있다. 어린 밀의 잎색깔은 식별하기 쉽고 콩의 비중은 측정하기 간단하므로 목표형질을 간접적으로 선발할 수 있다.
또한, 선발한 변이체의 유전자형을 알고자 할 때에는 호대검정(後代檢定; progeny test)을 한다. 이 방법은 변이체의 후대를 전개하여 형질의 분리 여부를 보고 동형접합체 또는 이형접합체를 판단한다.
최근에는 DNA 표지(marker)를 이용하는 분자표지이용선발(分子標識利用 選拔; marker-assisted selection, MAS) 기술이 개발되었다, 이 방법은 목표형질의 유전자와 연관된 분자표지를 선발하는 것으로, 포장에서 내냉성 검정이나 내병성 검정을 하지 않아도 된다.
2-3-2 생식
(1) 생식방법
①유성생식
작물은 종자 또는 영양체로 번식한다. 종자번식작물의 생식방법에는 유성생식(有性生殖; sexual reproduction)과 아포믹시스(apomixis)가 있고 영양번식작물은 무성생식(無性生殖; asexual reproduction)을 한다.
종자번식작물의 유성생식은 생식모세포(배낭모세포, 화분모세포, 2n)가 감수분열을 하여 암수 배우자(配偶子; gamete, n)를 만들고, 수정에 의하여 접합자(接合子; zygote, 2n)를 형성한다. 접합자는 개체발생을 하여 식물체로 자라며, 성장한 식물체를 포자체(胞子後; sporophyte. 2n)라고 한다. 암배우자(卵細胞)가 들어 있는 배낭과 수배우자(精細胞)가 포함된 화분(花粉)을 배우체(配偶體; gametophyte,n)라고 한다.
따라서, 유성생식은 하는 작문의 생활사(life cycle)는 배우체(n)세대와 포자체(2n) 세대가 번갈아 나타나는데, 이를 세대교번(世代交番; alternation of generation)이라고 한다. 포자체세대는 감수분열을 거쳐 배우체세대로 넘어가고, 배우체세대는 수정을 통해 포자체세대로 바뀌는데. 감수분열과 수정과정에서 유전변이가 생긴다.
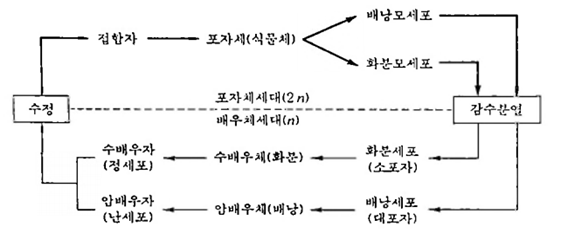
같은 개체에서 형성된 암배우자와 수배우자가 수정하는 것을 자가수정(自家受精; self-fertilization), 서로 다른 개체에서 생긴 암수 배우자 간에 수정이 이루어지는 것 을 타가수정(他家受精; cross-fertilizaion)이라고 한다. 그리고 자가수정을 하는 유성생식을 자식(自殖; selfing), 타가수정을 하는 유성생식을 타식(他殖; outcrossing)이라고 한다.
유성생식을 하는 종자번식작물은 주로 자식에 의하여 번식하는 자식성 작물(自殖 性作物)과 타식으로 번식하는 타식성 작물(他殖性作物)로 구분된다. 자식성 작물은 세대가 진전함에 따라 개체의 유전자형이 동형접합체로 된다. 그러나 타식성 작물은 세대를 진전하더라도 개제의 유전자형은 이형접합체로 남는다. 따라서, 육종에서는 자식성 작물과 타식성 작물의 육종방법이 달라진다.
1) 자식성 작물 : 벼, 밀, 보리, 콩, 완두, 담배, 토마토, 가지, 참깨, 복숭아나무
2) 타식성 작물 : 옥수수, 호밀, 메밀, 딸기, 양파, 마늘, 시금치, 호프, 아스파라거스
② 아포믹시스
아포믹시스(apomixis)란 'mix 가 없는 생식'을 뜻한다. 아포믹시스는 수정과정을 거치지 않고 배(胚)가 만들어져 종자를 형성하기 때문에 무수정종자형성(無受精裡子形 成) 또는 무수정생식(無受精生殖)이라고도 한다. 아포믹시스는 배를 만드는 세포에 따라 부정배형성 • 무포자생식 • 복상포자생식 • 위수정생식 • 웅성단위생식 등으로 나누어진다.
부정배형성(不定胚形成; adventitious embryony)은 배낭(胚襄)을 만들지 않고 포자체의 조직세포가 직접 배를 형성하며, 밀감의 주심 배가 대표적이다.
무포자생식(無胞子生殖; apospory)은 배낭을 만들지만 배낭의조직세포가 배를 형성하며, 부추, 파, 등에서 발견되었다. 복상포자생식(複相胞子生殖; diplospory)은 배낭모세포가 감수분열을 못하거나 비정상적인 분열을 하여 배를 만들며, 볏과, 국화과에서 나타난다.
위수정생식(僞受精生殖; pseudogamy)은 수분(受粉)의 자극은 받아 난세포가 배로 발달하는 것으로, 담배 • 목화 • 벼 • 밀 • 보리 등에서 나타난다. 위수정생식에 의하여 종자가 생기는 것은 위잡종(僞雜敏 false hybrid)이라 하고, 주로 종 • 속간 교배에서 나타난다. 웅성단위생식(雄性單爲生殖; male parthenogenesis)은 정세포 단독으로 분열하여 배를 만들며, 달맞이꽂 • 진달래 등에서 발견되었다.
아포믹시스에 의하여 생긴 종자는 수정을 거친 것이 아니므로 종자 형태를 가진 영양계라 할 수 있고, 다음 세대에 유전분리가 일어나지 않기 때문에 종자번식 작물의 우량한 아포믹시스는 영양번식작물의 영양계와 똑같이 곧바로 신품종이 된다.
③ 무성생식
무성생식은 생식기관이 아닌 잎 • 줄기 • 뿌리 등 영양체로부터 새로운 개체가 발생하며, 이를 영양번식(營養繁殖; vegetative propagation) 이라고 한다.
영양번식을 하면 유전적으로 동일한 특성을 나타내기 때문에 우량품종을 계속 증식하여 사용할 수 있다. 또한, 영양번식된 어린 식물은 모식물의 성숙한 조직에서 유래하였으므로 종자에서 발생한 어린 식물보다 강하다는 이점이 있다.
(2) 배우자형성
① 체세포분열과 감수분열
유성생식을 하는 작물은 체세포분열(유사분열)을 통해 개세로 성장하고 생식세포의 감수분열로 배우자를 만들어 생식을 한다. 생식모세포의 감수분열에 의하여 딸세포〔娘細胞; daughter cell)가 생기고 배우자로 되는 과정을 배우자형성(配偶子形成; gametogenesis)이라고 한다.
㉠ 체세포분열
체세포분열〔価細胞分裂. 유사분열(有絲分裂); mitosis〕은 하나의 체세포가 2개의 딸세포로 되며, 체세포분열은 일정한 세포주기(細胞週期; cell cycle)를 가지고 반복 적으로 일어난다.
세포주기는 G1기一S기—G2기 一 M기의 순서로 진행된다. G1 기는 딸세포가 성장하는 시기이고 S기는 DNA 합성으로 염색체(chromosome)가 복제되어 자매염색분체(姉妹染色分體; sister c&gmadd)를 만든다. G2기는 체세포분열을 준비하는 성장기이고 M기는 체세포분열에 의하여 딸세포를 형성한다. G1 기의 딸세포 중 일부가 세포분화를 하여 조직으로 발달한다.
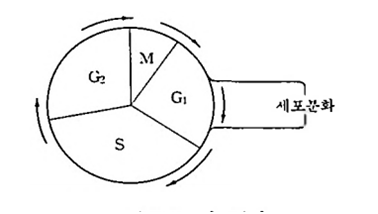
체세포분열은 전기 • 중기 • 후기 및 말기로 구분할 수 있다. 전기(前期; prophase)에는 염색사가 압축 • 포장되어 염색체 구조로 되며, 인과 핵막이 소실된다. 중기(中期; metaphase)에는 방추사가 염색체의 동원체에 부착하고 각 염색체는 적도판으로 이동한다. 후기(後期; anaphase)에는 자매염색분체가 분리되어 서로 반대방향으로 이동한다. 말기(末期; telophase)에는 핵막과 인이 다시 형성되고 세포질분열이 일어나 2개의 딸세포가 생긴다.
이와 같은 체세포분열은 체세포의 유전물질〈DNA)을 복제하여 딸세포들에게 균등하게 분배하기 위한 것이다. 그리고 체세포분열은 마모된 세포를 새것으로 교체하여 정상적인 기능을 수행하도록 하고 손상된 세포를 교체하여 상처를 치유하는 역할도 한다.
㉡ 감수분열
감수분열(減數分裂; meiosis)은 생식기관의 생식모세포(生殖母細胞; germ cell)에서만 이루어지며 연속적인 두 번의 분열을 거쳐 완성된다. 제 1감수분열은 생식모세포(2n)의 상동염색체(相同染色蚀; homologous chromosome)가 분리하여 반수체 딸세포(n)가 형성되는 감수분열(減數分裂; reductional division)이고, 제 2감수분열은 반수체 딸세포(n)의 자매염색분체가 분리하여 똑같은 반수체 딸세포(n)를 만드는 동형분열(同型分裂; equational division)이다. 감수분열의 과정은 전기 • 중기 • 후기 및 말기로 이루어진다.
제 1감수분열 전기는 세사기 — 대합기一 태사기— 이중기一이동기의 5단계로 나누어진다. 이 기간에는 염색사가 압축• 포장되어 염색체 구조를 이루며(細絲期), 상동염색체는 짝을 지어 2가염색체(bivalent)를 형성하고(對合期), 교차(交叉; crossing over)가 일어난다(太絲細). 그리고 2가염색체들은 적도판올 향하여 이동한다(移動期). 교차란 2가염색세 사이에 부분적 교환이 이루어지는 것을 말하며, 교차에 의하여 유전자 재조합이 일어난다. 교차로 인해 염색체가 꼬인 것과 같이 된 모양을 키아스마(chiasma, 복수 chiasmata)라고 한다.
2가염색체들은 적도판에 무작위로 배열한다(제 1감수분열 중기). 그리고 2가염색체의 두 상동염색체가 분리되어 서로 반대극을 향하여 이동하며(제1감수분열 후기), 새로운 핵막이 형성되어 반수체(n)인 2개의 딸세포가 생긴다(제 1감수분열 말기).
제 1 감수분열이 끝나면 극히 짧은 간기 (間期; interkinesis)를 거쳐 곧바로 제2감수분열이 시작되며, 간기에는 DNA 합성이 일어나지 않는다. 제 2감수분열은 반수체인 딸세포의 각 염색체에 있는 자매염색분체가 분리하며 체세포분열과 똑같이 진행 된다.
제1감수분열. 제2감수분열이 끝나면 1개의 생식모세포(2n)에서 4개의 반수체 딸세포(n)가 생기게 되며. 이들은 염색세 구성과 유전자형이 서로 다르다. 이는 제 1감수분열 과정에서 유전자재조합이 일어나고 염색체들이 재배치되었기 때문이며, 이는 유전변이(유전적 다양성)의 주된 원인이다.
감수분열은 유전변이의 원인이 될 뿐만 아니라 감수분열로 생긴 암배우자(n)와 수배우자(n)가 수정에 의하여 접합자(2n) 형성함으로써 생물종의 염색체수(2n)가 일정하게 유지된다. 또한, 감수분열 과정에서 상동염색체가 분리하기 때문에 Mendel의 유전법칙이 성립하게 된다.
② 화분과 배낭의 발달
식물의 생식기관은 암술과 수술이고 여기서 생식모세포의 감수분열이 일어나 배우자를 형성한다. 수배우자(정세포)는 수술의 화분(花粉, 수배우체)안에 있고 암배우자(난세포는 암술의 배낭(암배우체)안에 있다.
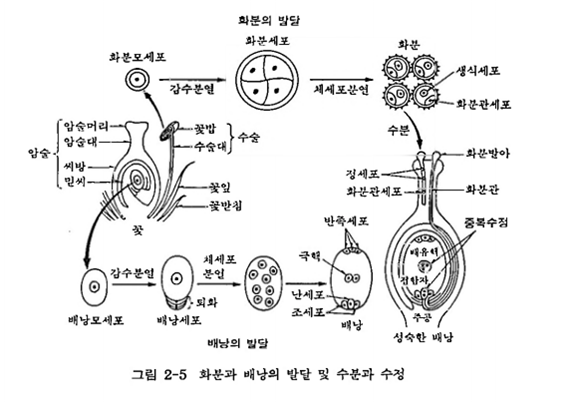
㉠ 화분
수술의 꽃밥 속에서 화분모세포 1개가 감수분열을 하면 4개의 반수체 화분세포가 형성된다. 화분세포는 두 번의 체세포분열이 일어나 화분(花粉, 꽂가루; pollen gn)으로 성숙하며, 각 화분에는 1개의 화분관세포(花粉管細胞; pollen tube cell)와 2개의 정세포(精細胞; sperm cell)가 있다. 화분관세포는 화분관으로 신장하여 정세포를 배낭까지 운반한다.
㉡ 배낭
암술의 씨방〔子房; ovary)속 밑씨〔胚珠; ovule)안에서 배낭모세포(胚涯母細胞; embryosac mother cell, EMC) 1개가 감수분열을 하면 4개의 반수체 배낭세포(大胞 子; megasporc)를 만든다. 그 중 3개는 퇴화하고 1개만 살아남아 세 번 체세포분열을 하여 배낭(胚囊; embryo sac)으로 성숙한다.
배낭에서 주공 쪽에는 난세포(卵細胞; egg cell) 1개와 조세포(助細胞; synergid) 2개가 있고, 주공의 반대쪽에 3개의 반족세포(反足細胞; antipodal cell)가 있으며 중앙에 극핵(極核; polar nucleus) 2개가 있다. 조세포와 반족세포는 나중에 퇴화한다. 주공은 화분관이 배낭으로 침투해 들어가는 통로이다.
(3) 수분, 수정 및 종자형성
① 수분
성숙한 화분은 꽃밥에서 터져나와 직접 또는 물, 바람, 곤충 등 매개체에 의하여 암술머리〔柱頭; stigma〕로 옮겨지는데, 이 과정을 수분(受粉; pollination)이라고 한다. 수분방법은 자가수분(自家受粉; self-poUination)과 타가수분(他家受粉; cross- pollination)으로 나누어진다. 자가수분은 한 개체의 화분과 암술 사이에 수분이 이루 어지는 것이고 타가수분은 서로 다른 개체간에 수분이 일어나는 것이다. 수분할 때 암술머리에는 여러 유전자형의 화분이 모이며, 수분이 성공하는 화분의 유전자형에 의하여 접합자의 유전자형이 달라진다.
자식성 작물 (自殖性作物)은 같은 꽃 속에 암술과 수술이 함께 있고(양성화), 암술과 수술의 성숙기가 같으며(자웅동숙). 자가불화합성을 나타내지 않기 때문에 자가수분에 유리하다. 또한, 자식성 작물은 꽃이 필 때 화기가 잘 열리지 않고, 화기가 열리기 전에 화분이 터지며, 암술머리의 위치가 자가수분에 적합하다. 자식성 작물의 타식률은 보통 4% 이하이다.
타식성작물(他殖褂伊豌)은 암술과 수술이 서로 다른 개체에서 생기거나(자웅이주 : 시금치• 삼• 호프• 아스파라거스), 수술이 먼저 성숙하거나(웅예선숙 : 옥수수• 딸기• 양파• 마늘) 또는 자식으로 종자를 형성할 수 없어(자가불화합성 : 호밀• 배추. 무, 메밀) 타가수분이 이루어지며 자식률이 5% 이하이다. 이러한 타식성 작물은 유전자형이 서로 다른 개체들 사이에 수분이 이루어지기 때문에 자식성 작물보다 유전변이가 더 크다.
② 수정
암술머리에 수분된 화분이 발아하면 화분관이 신장하며, 화분관을 따라 2개의 정세포가 주공을 통해 배낭 안으로 들어가 수정(受精; fertilization)을 하게 된다. 속씨식물〔피자식물(被子植物)〕은 2개의 정세포 중 하나가 난세포와 융합하여 접합자(2n)를 만들고 다른 하나는 극핵과 융합하여 배유핵(3n)을 형성하는데, 이 과정을 중복수정(気複受精; double fertilization)이라고 한다.
③ 종자형성
수정이 끝나면 배발생과 함께 밑씨가 성숙하여 종자(seed)로 되고 씨방이 발달하여 열매(fruit)를 형성한다. 종자의 배는 수정에 의하여 생겼으므로 한 세대가 진전한 것이다. 그러나 종피와 열매껍질〔果皮〕은 모체의 조직이다. 따라서, 종자에서 배와 종피는 유전적 조성이 다르다.
한편, 종자의 배유(3n)에 우성유전자의 표현형이 나타나는 것을 크세니아(xenia)라고 한다. 예컨대, 찰벼와 메벼를 교배하여 얻은 종자의 배유는 메벼로서, 이는 메벼 유전자 Wx의 작용에 의한 것이다. 사과 • 감 • 야자 등은 크세니아를 일으키는 유전자가 과일의 크기 • 빛깔 • 산도 등에도 영향을 끼치는데, 이를 메타크세니아(meta xenia)라고 한다.
수정으로 종자가 생성됨으로써 열매를 형성하는 것이 보통이다. 그러나 바나나, 감귤류, 포도, 중국감 등은 종자의 생성 없이 열매를 맺는 경우가 있는데, 이러한 현상을 단위결과(單爲桔果; parthenocarpy)라고 한다. 단위결과가 나타나는 것은 대부분 염색체 조성이 복잡하여 정상적인 배우자를 형성할 수 없기 때문이다. 단위결과는 자연적으로 일어나기도 하지만 다른 화분의 자극((예) 배추X양배추)이나 지베렐린과 같은 식물호르몬 또는 배수성((예) 건포도용 포도)을 이용하여 인위적으로 유발하기도 한다.
(4) 자가불화합성과 웅성불임성
① 자가불화합성
작물의 생식과정에서 환경적 • 유전적 원인에 의하여 종자를 만들지 못하는 것을 불임성(不稔性; sterility)이라고 한다. 유전적 불임성에는 자가불화합성(自家不和合 性; self-incompatibility)과 웅성불임성(雄性不稔性; male sterility)이 있으며, 이는 식물이 유전변이를 확대하기 위한 수단이다. 작물의 자가불화합성과 웅성불임성은 1대잡종(F1) 품종의 종자를 채종하는 데 이용된다.
자가불화합성은 암술과 화분의 기능이 정상적이나 자가수분으로 종자를 형성하지 못해 불임이 생긴다. 자가불화합성의 메커니즘은 암술머리에서 생성되는 특정 단백질(S-glycoprotein)이 화분의 특정 단백질(S-protein)을 인식하여 화합, 불화합을 결정하게 된다. 불화합이면 암술에서 생성되는 억제물질에 의하여 화분이 발아하지 못하고 발아해도 화분관이 신장할 수 없다.
자가불화합성은 S유전자좌의 복대립 유전자가 지배하며, 자가불화합성의 유전양식에는 배우체형과 포자체형이 있다. 배우체형 자가불화합성은 화분(n)의 유전자가 화합• 불화합을 결정하고, 포자체형 자가불화합성은 화분을 생산한 식물체(포자체, 2n)의 유전자형에 의하여 화합 • 불화합이 달라진다. 배추는 자가불화합성의 유전자형이 다른 자식계통을 혼식하여 1대잡종 종자를 채종한다.
② 웅성불임성
웅성불임성은 유전자작용에 의하여 아예 화분이 형성되지 않거나 화분이 제대로 발육하지 못하여 수정능력이 없기 때문에 종자를 만들지 못한다. 웅성불임성에는 핵 내 ms 유전자와 세포질의 미토콘드리아(mitochondria)DNA 가 관여한다.
핵내 유전자만 작용하는 웅성불임은 유전자웅성불임성(遣傳子雄性不稔性; genic male sterility, GMS)이라 하고, 세포질유전자만 관여하는 웅성불임은 세포질웅성불임성(細胞質雄性不稔性; cytoplasmic male sterility, CMS)이라 하며, 핵내 유전자와 세포질유전자의 상호작용에 의한 웅성불임은 세포질•유전자웅성불임성(細胞質•遺傳子雄性不稔性; cytoplasmic-genic male sterility, GGMS)이라고 한다.
세포질•유전자웅성불임성(CGMS)은 화분친의 임성회복유전자{稔性回復遗傳子; fer tility restoring gene, 用)에 의하여 임성이 회복된다. 따라서, 이 웅성불임계통(자방친)에다 임성회복유전자를 가진 계통(화분친)을 교배하여 1대잡종 종자를 채종한다. 세포질웅성불임성(CMS)은 화분친에 관계없이 불임이 되므로 양파처럼 영양기관을 이용하는 작물에서 1대잡종을 생산하는 데 이용된다.
유전자웅성불임성 중에는 온도• 일장• 지베렐린 등에 의하여 임성을 회복하는 환경감응형 웅성불임성(環境感應型雄性不稔性)이 있다. 벼의 온도감응형 유전자웅성불임성은 21~26℃ 에서 임성을 95% 이상 회복하므로 1대잡종 종자를 채종하는 데 이용할 수 있다.
'농업 공부 > 재배학 이론' 카테고리의 다른 글
| 재배학원론 - 2장 작물의 유전성(4) : 유전양식1 (0) | 2022.10.21 |
|---|---|
| 재배학원론 - 2장 작물의 유전성(3) : 염색체와 유전자 (0) | 2022.10.21 |
| 재배학원론 - 2장 작물의 유전성(1) : 품종 (0) | 2022.10.20 |
| 재배학원론 - 1장 재배개설(2) (0) | 2022.10.20 |
| 재배학원론 - 1장 재배개설(1) (0) | 2022.10.19 |




댓글